Chemical Signaling
Chemical Signaling
The primary means by which cell-to-cell communication, as well as homeostasis in multicellular organisms, occurs, involves protein receptors by receiving cells. Chemical signaling can involve small molecules (ligands) or large molecules (cell-surface signaling proteins).
Read More: What is signaling in biology?
Chemical signaling and orientation
The chemicals used in the bee signaling and orientation system can be divided into two groups. The first includes those that are secreted by the bees themselves, the second - natural odorous substances that are part of food and propolis, as well as the secretions of enemies of bees. Both of them are used by bees to coordinate many behavioral acts, the understanding of which is impossible without an analysis of the role of chemical signals.
Example:

- Direct signaling (also called juxtacrine signaling) involves communication between cells that are in direct contact with each other. This communication is often mediated by gap junctions in animal cells and plasmodesmata in plant cells.
- Autocrine signaling occurs when a ligand acts on the same cell that releases it.
- Paracrine signaling occurs when a ligand diffuses in a small area and only acts on neighboring cells. Neural signals are a specialized subset of paracrine signals, diffusing a very short distance within the synaptic cleft between adjacent neurons (more on neural communication in a future class session).
- Endocrine signals are hormones, which are carried throughout the organism’s body via the vascular system (in either plants or animals!) to act on cells that may be very far away from the cells which released the ligand (more on plant and animal hormones in future class sessions). Neuroendocrine signals are a specialized subcategory of endocrine signals and are released by neurons but that travel via the vascular system to act on cells that are far away.
- Pheromones are released into the environment to act on the cells in a different individual.
Example 2:
Signaling in single-celled organisms
- formation of biofilms
- attacking competitors
- luminescence (emitting light)
- activation of virulence genes (associated with pathogenicity, or the ability to cause an infection)
How does quorum sensing work?

In this section, we describe only some of the mechanisms of release of odorous substances by bees and the work of chemoreceptors.
Mechanisms Of Odorous Substances By Bees
The excretion of odorous substances by bees. Of the specialized odorous glands, Nasonov's gland has been well studied. It is found in worker bees and queen bees. The gland is located between the 5th and 6th dorsal segments of the abdomen. Worker bees protrude the gland, lifting up the abdomen. This is an external manifestation of the functioning of the gland.

Many researchers have noted that Nason's gland most often functions in worker bees when they visit food sources that do not have a smell. Bees also use this gland when they are engaged in the delivery of water, the smell of which can suppress or activate the functioning of the gland. Thus, J. Free and J. Williams (1970) noticed that 53-67% of the bees visiting the feeder with distilled water exhibited iron. When odorous substances were added to the water, the number of these bees decreased to 5-11%.
The secret of Nasonov's gland contains geraniol, citral, geranium, and nerolic acids. Of these chemical components, the cis- and trans-isomers of citral have the greatest attraction (attractiveness) for bees. The odorous substances secreted by the Nason gland are contained in the nectar and pollen of flowering plants visited by bees. This suggests that food influences the chemical structure of pheromones. From the standpoint of the foregoing, it is possible to explain the phenomenon known as "floral specialization", which is expressed in the fact that bees of different ecological populations prefer certain types of plants from among those that simultaneously bloom and produce nectar and pollen. Apparently, this feature of bees of different populations was formed in the course of their long conjugated evolution with plants. Hence, for the optimal functioning of the whole complex of physiological processes occurring in the bee colony, its members require nectar and pollen of certain plant species, characteristic of the conditions in which the ecological population was formed.
The uterus uses Nasonov's gland when flying out of the home. The odorous secretions of the gland serve as a guide for the bees accompanying the queen during the swarming period. To detect the queen bees also use the secretion of her mandibular glands. Their secretions are part of a complex of odorous substances that attract drones to the uterus during the mating season. They are also excited or attracted by secretions of accumulations of subepidermal cells located on the 2nd, 3rd, and 4th abdominal tergites (M. Renner, M. Baumann, 1964).
Repellents are secreted, as noted above, by the poisonous gland. Probably, isoamyl acetate is among the repellent substances she secretes. The excretion of the mandibular glands of worker bees also appears to contain repellents. They, according to the observations of D. Simpson (1966), have a deterrent effect on worker bees.
Chemoreceptors
Various chemicals are perceived by chemoreceptors, which are functionally divided into olfactory (olfactory) and taste (contact). Olfactory react to odorous substances, the molecules of which are present in the air. Contact - on substances dissolved in water.
Olfactory receptors are located on the eight distal segments of the antennae. This is indicated by the results of electrophysiological and behavioral experiments. Bees with eight distal antennae amputated do not respond to odorous substances. G. Demirchoglyan and V. Neprimyan (1971) by recording electro antenna grams during sequential amputation of antenna segments established that chemoreceptors located on the sixth, seventh and eighth segments have the greatest sensitivity.
Morphologically and anatomically, seven types of sensilla are distinguished on the antennae. Of these, so far only placoid sensilla can be confidently attributed to olfactory receptors. Different groups of these sensilla are specialized for the perception of certain chemicals. However, the fact that bees distinguish a wide variety of odorous substances is probably due to the fact that the working ranges of various receptors overlap. The results of the work of E. Veireshi (1971) speak about the incredibly high working range of the chemoreceptor apparatus of bees. In his experiments, out of 1816 paired combinations of odors presented to him, bees distinguished 95.5%.
Placoid sensilla are oval plates. Their diameter reaches 12-14 microns. The plates are permeated with pores, the diameter of which ranges from 0.01 to 0.03 microns. Each placoid sensilla contains 12 to 18 receptor cells.
The number of placoid sensilla on the antennae of bees, queens, and drones is not the same. Worker bees have 3600 to 6000 sensilla on each antenna. The uterus has about 3,000, and the drone has 30,000 placoid sensilla. An increase in the number of sensilla directly correlates with the number of afferent impulses arriving in the central nervous system with the same olfactory stimulation. This and the above information about the different number of placoid sensilla in different representatives of the bee family allows us to assert that the sense of smell is most developed in drones.
It is generally accepted that the sense of smell of bees is more developed than that of humans. This is true only for certain odorous substances. For a number of substances, the sense of smell of bees is at the same level as that of a person, and for some, it is much lower than it.
The contact chemoreceptors of the bee mainly control the suitability of nectar, water and other substances for nutrition. These receptors are found on the mouthparts (labial palps), distal segments of the legs (tarsi) and antennae. Antenna contact chemoreceptors are represented by trichoid and basiconic sensilla.
Taste buds stimulate positive or negative reactions of bees to various substances. So, bitter and sour substances cause a negative reaction in bees. A positive food reaction is stimulated by sucrose, glucose, maltose, fructose, trehalose, melecytose, fucose, and mitylglucoside and inositol. The attractiveness of these substances varies. For oral receptors, the attractiveness of glucose is equal to the attractiveness of maltose, fructose is equal to glucose. Glucose attracts bees more than fucose, and fucose is superior to galactose.
The labial receptors of bees are characterized by a relatively low sensitivity to the concentration of carbohydrates. The threshold sugar concentration for these receptors is at the level of 0.062-0.125 M, which is much lower than the sensitivity of the contact receptors located on the antennas. The latter react to a solution of sucrose, the concentration of which is only 0.01-0.07 M. The low sensitivity of the labial receptors to sugars is explained by the fact that the bees have adapted to consume highly concentrated solutions of sugars. The positive reaction of bees to sugar solutions is expressed in the fact that they extend their proboscis and begin to take this carbohydrate food. This reaction is stimulated by contact receptors located on the legs and antennae. The threshold concentration for the same sugars can vary depending on the physiological state of the bees by more than ten times. In hungry insects, ceteris paribus, the taste thresholds are much lower than in satiated insects.
The sensitivity of the taste buds of bees, as well as olfactory ones, for some substances is close to the taste sensitivity of a person, for others it is very different from it. For example, a bee begins to distinguish distilled water from a sodium chloride solution when its concentration reaches 0.24 M. Human taste buds detect the presence of sodium chloride at a concentration of 0.09 M. The sensitivity thresholds for hydrochloric acid in bees and humans are the same. They are 0.001 M. In terms of sensitivity to quinine, the taste analyzer of a person is about 100 times higher than that of a bee. It reacts to the presence of quinine when its concentration is only 1-10-5. The given sensitivity values indicate that it varies greatly depending on the chemical structure of the substance.
Chemoorientation
The location of olfactory receptors on mobile organs - antennas - makes it possible to determine the position of an odorous target for sitting and flying bees. The role of antennas in olfactory orientation is shown by the works of X. Martin and M. Lindauer (1963, 1964, 1965). They conducted a series of special experiments in which the bees were accustomed to visit flavored top dressing. The ability of the bees to navigate by the smell, to which they developed a preliminary conditioned reflex, was assessed by the direction of movement chosen by the insects in the Y-shaped maze. It turned out that intact bees found the way to the goal the fastest. During the search, they moved apart the antennas and moved (chanted) them. After amputation of one of the antennas, the bee found the target much more slowly. A bee with fixed antennae moved even more slowly towards the target. This indicates the importance of oscillatory movements of the antennas during the search. This is confirmed by the results of an experiment in which one of the antennas was removed and the other was fixed on the head. Such a bee moved towards the target along a zigzag path.
The difference in the concentration of an odorous substance, necessary for the bee to recognize the direction to it, was determined in a series of experiments by X. Martin. To do this, he put glass capillaries on the antennas, in the distal ends of which an aromatic substance of various concentrations was placed. According to the chemotropotactic reaction (turn to the action of a chemical stimulus) of bees, the necessary ratio of the concentration of an odorous substance acting on different antennas was determined. In these experiments, the bee turned when the concentration ratio of odorous substances reached 2.5. Therefore, a 2.5-fold concentration gradient of the odorous substance is sufficient for the bee to be able to detect the direction towards the odorous target.
Strategies
Three strategies of chemical signaling are the use of hormones, local chemical mediators, and neurotransmitters.
Chemical transmission in synapses is based on the same principles as chemical signaling using water-soluble hormones. In both cases, the cell releases a mediator substance that acts on another cell or group of cells by binding to membrane receptor proteins. However, unlike the hormone, the chemical messenger in the synapse is the neurotransmitter- works only at very short distances.
The author of the Introduction to Chemical Ecology, Michel Barbier, is a prominent specialist in the field of bioorganic chemistry. Over the past twenty years, his focus has been chemical signaling in insects, sterol metabolism, marine invertebrate chemistry, phytopathology, and other topics. The breadth of scientific interests, happily combined with the art of presentation, helped him solve a very difficult task in a concise and vivid form to talk about the basic concepts of chemical ecology.
Therefore, the work of genes is indeed regulated by chemical signaling.
Three different ways of intercellular chemical signaling
There is, therefore, chemical signaling between genes, carried out through the mediation of the cytoplasm.
Thus, and mathematically, there is a far-reaching analogy between electrical and chemical signaling, and that is why electrical simulation of chemical reactions is possible.
Intercellular chemical signaling
Three types of chemical signaling are known 1) most body cells secrete one or more signaling substances, which serve as local chemical mediators since they are destroyed or absorbed so quickly that they affect only the cells of the immediate environment 2) specialized endocrine cells secrete hormones that are carried by the blood and affect target cells, and the latter can be located in various parts of the body 3) nerve cells form specialized contacts with their target cells chemical synapses ) and secrete chemicals that act only at very short distances - neurotransmitters that affect (in each synapse) only one target cell.
Endocrine cells and nerve cells are specialized for different types of chemical signaling Intercellular chemical signaling
Intercellular
chemical signaling 249 and nerve cells are specially designed for chemical signaling, they jointly coordinate the various activities of the billions of cells that make up the body of a higher animal.
they transmit information much faster than endocrine ones, since, for signal transmission over long distances they do not need diffusion and blood flow, the signal is quickly transmitted along with the nerve fiber by electrical impulses. It is only at the nerve endings where the neurotransmitter is released that these impulses are converted into chemical signals. The neurotransmitter reaches the target cell by diffusion at a microscopically small distance, which takes less than a millisecond (Fig. 13-2). While hormones are highly diluted in the circulation and must be able to act at extremely low concentrations (usually <10 M), the dilution of neurotransmitters is negligible and their concentration in certain areas of target cells can be high. For example, the concentration of the neurotransmitter acetylcholine in the synaptic cleft of the neuromuscular junction can be as high as 5 M. However, in other respects, the mechanisms of chemical signaling by hormones and neurotransmitters are broadly similar, and many of the signaling molecules used by endocrine cells are also used nerve cells (neurons).
Many types of chemical signaling between the cells of multicellular animals could arise from the adaptations by which unicellular organisms respond to chemical changes in the environment . Indeed, as already mentioned, some intracellular mediators are used by organisms of both types. The most studied reactions of unicellular organisms to external signals include chemotaxis phenomena, in which cell movement is directed towards or away from the source of a particular substance. Chemotaxis of eukaryotic cells is discussed in Sec. 14.3.2 on the example of the cellular stage of the slime mold Dictyostelium dis oideum and in sec. 11.6.4 on the example of human neutrophils. We will end this chapter with a description of bacterial chemotaxis, which, largely due to advances in genetic analysis, illustrates especially clearly and elegantly the central role of adaggation in responses to chemical signals. [c.385]
Chemical signaling also plays an important role in the processes of animal development, often determining the time and type of differentiation of certain cells. In such cases, the effect of the chemical signal is usually slow and long lasting. For example, during puberty, ovarian cells begin to secrete the steroid female sex hormone estradiol in large quantities. This hormone causes changes in many cells in various parts of the body, which eventually leads to the development of female secondary sexual characteristics, such as breast enlargement. If estradiol secretion stops, these effects gradually disappear, but some reactions caused by steroid sex hormones at a very early stage of development mammals are irreversible (see the end of Table 13.1). Similarly, a tenfold increase in the concentration of thyroid hormone in the blood causes a series of radical and irreversible changes, leading to the transformation of a tadpole into a frog.
Chemical transmission in synapses is based on the same principles as chemical signaling with the help of water-soluble hormones. In both cases, the cell, through exocytosis, releases a chemical agent into the intercellular space - an intermediary, which then acts on another cell or group of cells by binding to membrane receptor proteins. In the synapse, such an intermediary, a keyrokdiator, passes by diffusion from its source to the target a distance of only a fraction of a micrometer, while the hormone can mix with blood flow to distant parts of the body. However, this is not a fundamental difference. Some substances perform dual duties - they act both as circulating hormones released by cells of the endocrine glands and as neurotransmitters released by nerve endings. Moreover, some clepsies are typical in their form and ability to conduct impulses
Apparently, it was insects that, during their evolution, we're able to develop the most sophisticated system of chemical signaling, which plays a huge role in their life. Indeed, with the help of pheromones, they are able to transmit information about the presence of individuals of the same or another species (recognition signals like army identification signals friend or foe), the location of a male or female (sexual attractants), the approach of danger ( alarm pheromones ), the presence of a source food and the route to it (aggregation pheromones and trace markers ) and much more.
In social insects, as well as in insects that temporarily form large communities, the action of pheromones must be regulated very finely and take into account the situation. Usually, multicomponent systems are used here for chemical signaling, in the formation of which several glands can participate. It can be assumed that the meaning of the signal changes depending on the intensity with which one or another component of the signal mixture is produced by the corresponding gland. However, this aspect of intraspecific interactions due to its complexity, very little has been studied. Ants A anihomyops laviger are known to have two glands that produce alarm pheromones. Dufour 's abdominal glands contain hydrocarbons (161, 162 and 164) and methyl ketones (163 and 165), while the mandibular glands secrete a number of terpenoids , including norcitronellal (166), citronellol (167), citronellal (168), neral (169 ) and geranial - (170) with the exception of pentadecane (164) and pentadecanone -2 (165), all of the listed substances are effective alarm pheromones[413]. A disturbed ant of this species releases a small amount of alarm pheromones from both glands , which immediately cause anxiety in nearby tribesmen. Attracted by pheromones, they rush to the alarm ant, and at the same time release the same pheromones themselves. From ant to ant, this signal spreads with the speed of a chain reaction , and soon the whole anthill comes to combat readiness. Alarm pheromones are widespread in insects.
Chemical signaling in the body is extraordinarily complex, but gradually it is possible to understand it. Here, the sources of signals and their progenitors are equally important. The craftsmanship of nature is magnificent, it created not only the most complex self-regulating systems that work smoothly, but also the human brain, which turns out to be able to cognize the essence of this self-regulation, to cognize itself.
One way or another, it has now become clear that visual perception occurs by converting a light signal into a molecular, chemical one. The information brought by a light quantum is recoded into chemical information through retinal isomerization. In turn, this signal causes a conformational rearrangement of the protein. We do not yet know how such a rewiring creates a nerve impulse , and how nerve impulses produce a sensation of light and color in the brain . But the basis of vision is again chemical signaling.
Insects and some lower plants in the process of evolution were able to develop chemical signaling systems between individuals that play a huge role in their lives. Chemical mediatorsin the transfer of information between individuals of the same species, compounds belonging to different classes and differing in structure and functions act. For such compounds, the term pheromones or exohormones, is proposed, in contrast to the endohormones which regulate the functions of cells and organs within a given organism.
Experiments in nature have shown that chemical signaling determines not only the abundance of a species but its quality, the genetic structure of its population. This means that the chemical code ultimately determines through the number of dominant species and the structure of the biocenosis, and the structure of populations. its constituent species, and their abundance. Having mastered the chemical code of regulation of biocenosis and population processes, we will be able to control the most complex natural processes without fear of adverse side effects.
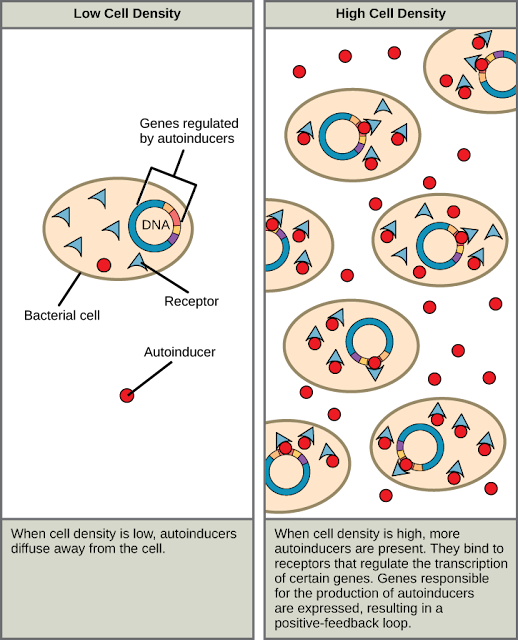
Chemical signaling in Dictyostelium is the most studied example of chemotaxis in eukaryotic cells. In response to a lack of food, amoeba begins to synthesize and release cyclic AMP (cAMP) in successive pulses. For unknown reasons, some cells become centers of aggregation. The cAMP secreted by them binds to specific receptors on the surface of other starving amoebae and orients their movement towards the cAMP source. This behavior can be directly demonstrated by applying a very small amount of cAMP with a micropipette to any part of the surface of a starving amoeba. The response to this exposure will be the immediate formation of pseudopodia growing towards the micropipette. Under normal conditions, the pseudopodia would attach itself to the surface the cell is on and pull the cell along in the same direction.
Comments
Post a Comment